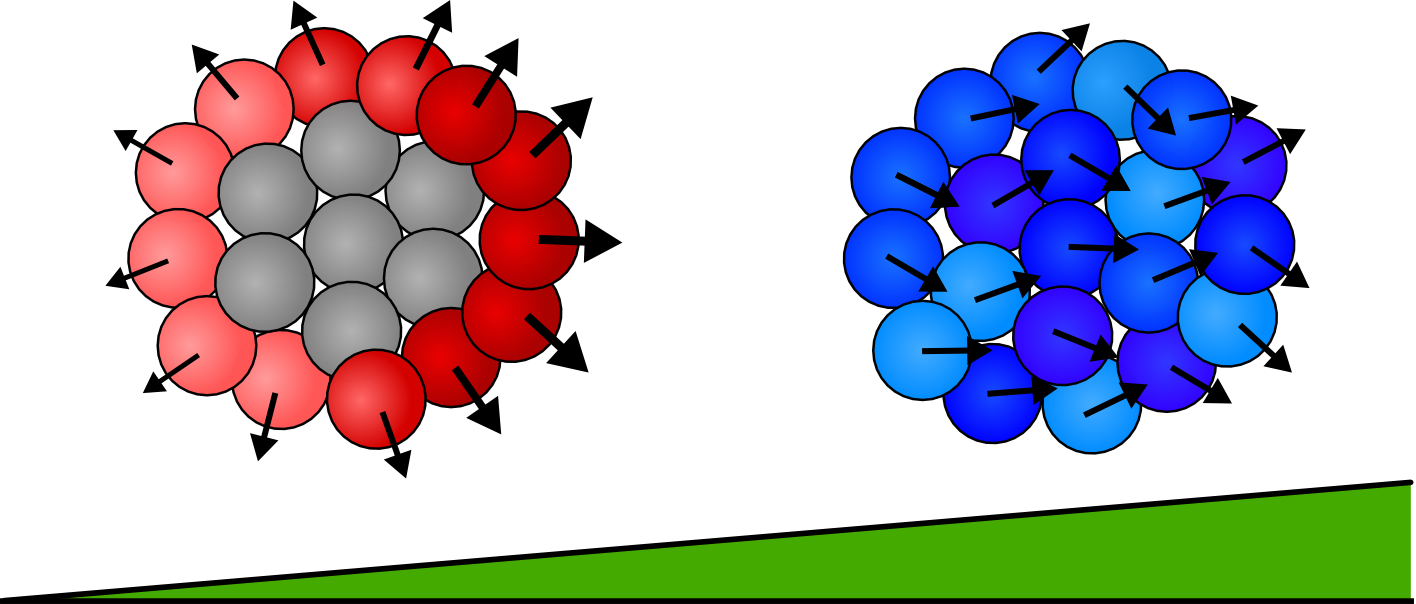
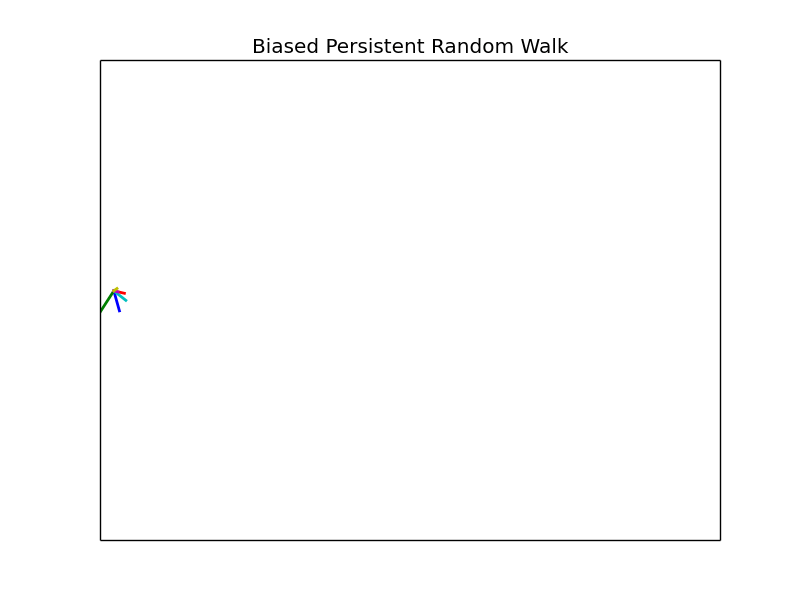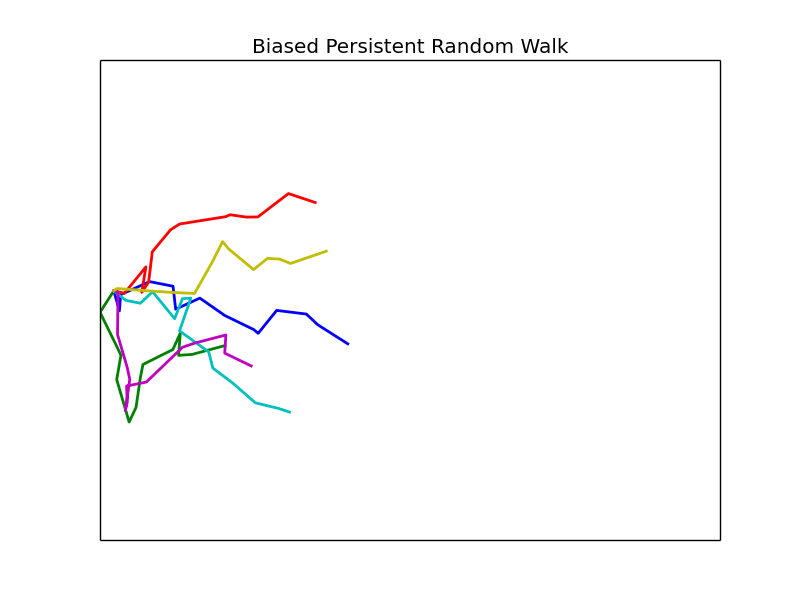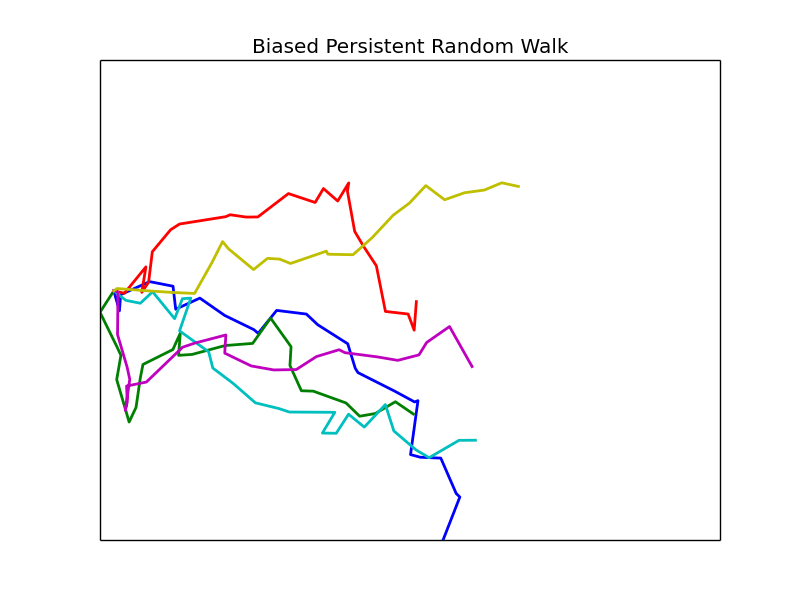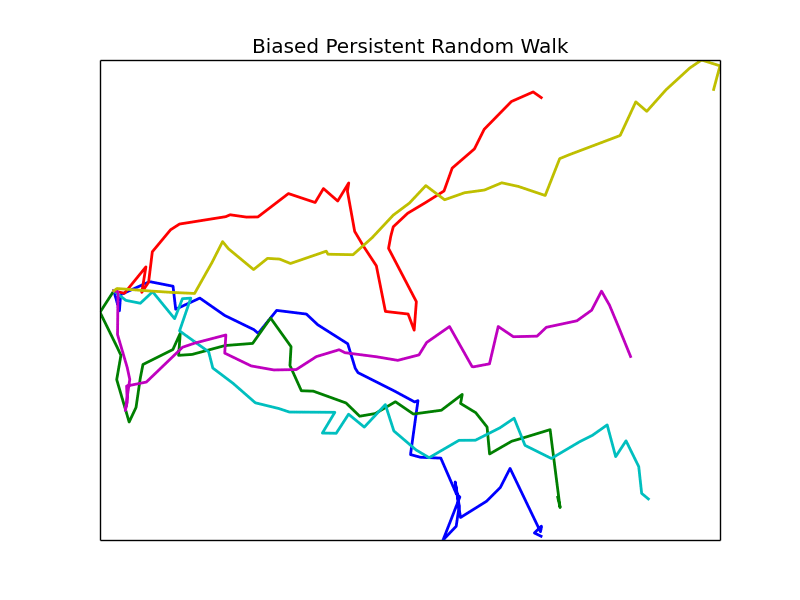
Phase Space
We vary the parameters \(\beta\) and \(\epsilon\) and observe the resulting mean first passage time. Other parameters are left fixed unless otherwise noted, all plots are of groups of 4 cells.
No Perimeter Energy Constraint
Read More
Cell Sensing, Implementation in CPM Code
Overview
Cells in a group communicate with one another in order to sense a gradient. We use a minimal adaptive model based on local excitation and global inhibition (LEGI). The network looks...
Read More
Simulation Parameters:
- Each cell has a relaxed area of 27 lattice sites.
- Cells are initialized as
3x3x3 cubes.
- Each simulation runs for 300 Monte Carlo (MC) time steps.
- Statistics in each case are gathered...
Read More
1 Cell
Resting Area of 4 Pixels
The simulation ran for 100 different instances each for a maximum of 60 Monte Carlo (MC) time steps.
From the simulations we were able to get statistics on the mean square...
Read More
Adaptation
We will continue our discussion on adaptation from the last lecture. We want to answer the question of what causes the sharp decrease (response) in activity \(A\) due to an increase in ligand concentration \(l\).
Response Curve
...
Read More
Bacteria Forward Motion & Tumbling
As previously discussed, most bacteria use flagella in order to achieve forward motion. Bacteria then tumble in order to change direction. Experiments have shown that bacteria in environments of high concentration of some attractant...
Read More
Cells as Random Walkers
Model
The goal is to simulate a connected, one-dimensional chain of freely diffusing cells. We model the chain of cells by only worrying ourselves with the edges of each cell since we are interested...
Read More
Competance in Bacteria
Competance is a state in which bacteria can take in DNA from the environment and add it to their own. It is a desperate action in attempt to speed up evolution in order to survive.
We...
Read More
Repressilator
A repressilator is a feedback loop in which each gene suppresses the next one. The following diagram shows an implementation of a repressilator.

Questions about the repressilator
- When does the repressilator oscillate...
Read More
Feedforward Loops
Recall the different types of feedforward loops. Last lecture we discussed the behavior of the C1 loop. The advantage of the C1 loop is that it protects Z from turning on due to fluctuations in X. This...
Read More
Network Motifs
We have already examined the one node motif of auto-regulation, let’s now explore 3 node motifs.
Let \(n\) be the number of nodes and \(g\) be the number of edges. For a 3 node motif \(n=3\) and...
Read More
Cell Volume as a function of Alpha
Model
\[\beta H_s = \sum_{j=1}^N \left[\alpha\left(V_j/V_0-1\right)^2-\sigma_j\gamma\right]\]
A simple check that the simulations are working is to see how the cell volume \(V_j\) changes as \(\alpha\) changes.
We expect that for large...
Read More
Autoregulation (continued)
Can we say something quantitative about…
- Noise reduction for autorepression.
- Switching time for auto-activation.
Last lecture we came up with an expression for the steady-state probability distribution for an autoregulating protein, but the expression was...
Read More
Autoregulation
We will continue our discussion of autoregulating genetic networks. Specifically, we will continue talking about autorepression.
Autorepression
Recall that there are two advantages of autorepression.
- Reduces response time.
- Buffers fluctuations.
Noise Reduction (buffer fluctuations)
...
Read More
Network Motifs
In this lecture we will broaden our scope to a whole genetic network.
We define a motif as a pattern that is over-represented in a network. A pattern is considered to be over-represented by comparing it with...
Read More
Noise in Gene Activation
We will continue our discussion of noise in gene activation from the previous lecture.
Last lecture we made a simple model for gene activation and the intrinsic noise in the system. Next we...
Read More
Gene Activation
In the previous lecture we discussed a simple model of protein production with an activator.
\[\text{X}+\text{D} \rightleftharpoons^{k_+}_{k_-} \text{C} \xrightarrow{k}\text{Y}+\text{X}+\text{D}\]
Now lets consider a more involved model.
Additional Considerations
X has its own dynamics
\[\text{D} \xrightarrow{g}\text{D}+\text{X}...
Read More
Gene Networks
Starting with this lecture we will shift our attention in this course. Instead of focusing on physical characteristics of biological systems we will focus on functional control of biological systems.
Here is a simple example of a...
Read More
The Algorithm
There are three cases that the code handles.
- Single cell diffusion
- Edge cell diffusion
- General case diffusion
The single cell case is treated as a simple random walk. The edge cases and the...
Read More
Double Headed Motors
Experimentally, the speed and the force on a loaded motor were calculated.
\[F_\text{load}=6.5 \ \text{pN} \\ v_\text{motor}\approx 150 \ \text{nm/s}\]
Unloaded Motor
In the case of an unloaded motor there is no force.
\[v \approx...
Read More
Molecular Motors

amazing picture. (source)
All motors rectify thermal motion. To make a molecular motor its potential energy \(U(x)\) needs to have one of the two following properties.
- \(U(x)\) is modulated in...
Read More
Dynamic Instability of the Microtubule
What is the timescale of fluctuations? (\(\tau\))
A good guess would be that it depends only on the two rates which we are aware of - the attach and detach rates.
\[\tau \approx...
Read More
Simulation Set-up
- Reflective boundary on the left.
- Absorbing boundary on the right.
- Cells are always connected, cells may stretch and contract.
- The number of cells is varied.
- The amount by which a cell can stretch is varied....
Read More
1 Cell Diffusion with No State Switching
$$ P(x|t) $$
Prediction
The probability density for cell location given a time \(t\) should look like a normal distribution. As time increases this prediction becomes less accurate since more...
Read More
Dynamics: Filament Polymerization
Polymerization at Both Ends (cont.)

Actin
Unlike microtubules, actin does not have the same critical concentration for both ends. The “\(+\)” end is the side where the filament grows and...
Read More
Flagellum Propulsion
What is the efficiency of the flagellum?

We are interested in calculating the efficiency of the flagellum as a means of propulsion for the bacteria. Efficiency is defined as the following:
\[\mathcal{E}...
Read More
Bacteria Forward Motion
How do bacteria achieve forward motion?
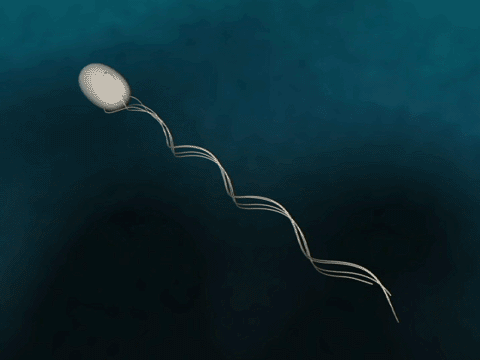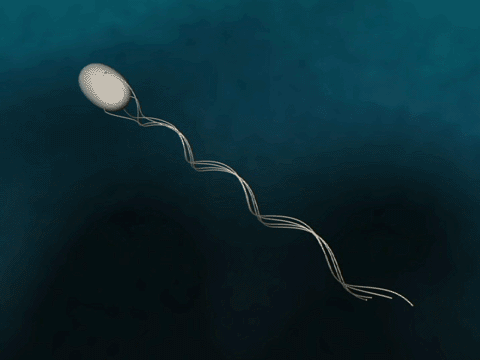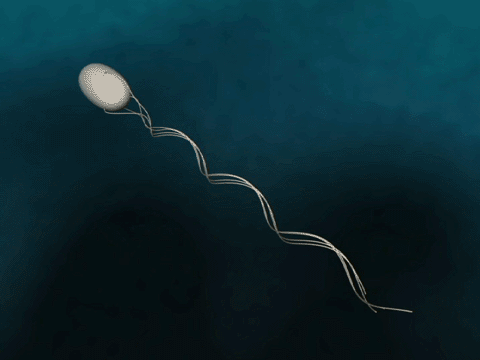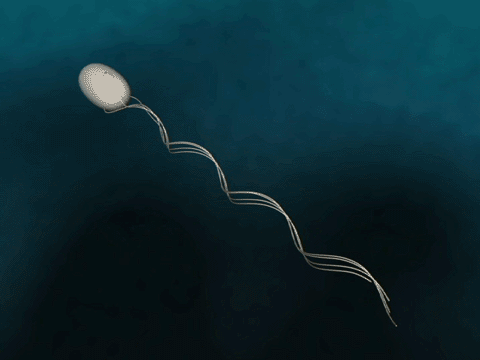 source
source
Bacteria have flagella that drive their propulsion. Bacteria flagella move in a corkscrew pattern. Flagella do not flap back and forth like a paddle....
Read More
We will start be continuing in our answer to the question:
What shapes emerge from beinding energy considerations alone?
Pancake
A pancake is a cylinder with rounded sides.

Let the rounded sides have height...
Read More
Lipid Geometry
What determines the shape of grouped lipids?

Micelle
A good guess is that conical shaped lipids are more likely to form micelles. Let a cone-shaped lipid have a length \(l\), circular...
Read More
Self-Avoiding Chain
- A self-avoiding chain is a freely jointed chain the does not cross over itself.
- One of the first scientists to work on this problem is Flory.
- We want to quantify how the...
Read More
Elasticity (Force-Extension Relations)
Consider a polymer where one end is fixed while the other end is free.

Now we apply a force, \(\vec{f}\), to the free...
Read More
Polymers
- In previous lectures we discussed Freely Jointed Chains (FJC) and Freely Rotating Chains (FRC) and derived some properties relating to them.
- We calculated some of the following properties for the FJC and FRC in 3D. Now we...
Read More